







Urban Allergy: Of Mice and Humans
Where patients live makes a difference.
BY MARK B. ABELSON, M.D., AND LAUREN LILYESTROM
Aesop's fable of the city mouse and the country mouse, where the simple country mouse visits his urban friend, only to be chased from his fancy meal by dogs, concludes with the moral of "better beans and bacon in peace than cakes and ale in fear." The country mouse in Aesop's fable thought that urban life was a little more than he bargained for. For many people, the hustle and chaos of cities provide excitement, entertainment and employment; however, cities are also taxing on our immune systems.
Our dependence on industrialization and commerce has made rural living an inconvenient — if not impossible — way to obtain relief of itchy, red eyes and runny noses. More is involved in the epidemiology of allergy than simply hygiene and urban pollution. This article will discuss some of the factors that are contributing to the increasing incidence of eye allergies. Given the abundance of allergic mediators, it is not surprising that many genes are believed to be involved in the allergic response.
Environmental Correlations
The incidence of allergy appears to be increasing, and approximately 20% of the population endures the signs and symptoms of allergic conjunctivitis.1 Many epidemiological studies have reported a dichotomy between the prevalence of allergy in the country vs. that in the city. The prevailing hypothesis to explain this phenomenon is that allergens and soil microbiota that are encountered on a daily basis on the farm shift the immune system from the neonatal T helper (Th2)-favoring system to a Th1-favoring system, thereby preventing the allergic response.
The developing fetus is essentially a foreign graft to the maternal immune system. Accordingly, the maternal immune response to the fetus is Th1-mediated, a response that is also associated with graft and transplant rejection.2 To counter this effect, the fetus maintains a Th2-biased response to direct the maternal system away from Th1.3 It is not until parturition that the maternal Th1 response strengthens.4 Early stimulation of the immune system by pathogens and allergens is thought to shift the immune system's focus away from Th2-mediated responses. Even consistent childhood exposure to the farming environment, however, does not always protect against allergies upon relocation to the city.5 Furthermore, the incidence of asthma and atopy among homeless children — who presumably have high exposure to allergens and microbiota — is significantly higher than that which is usually observed amongst urbanites.6
Pollution — specifically the oxidative properties of exhaust fumes — is therefore believed to interact with genes and the physiological processes of allergy. The number of biological and environmental factors that commingle to produce the allergic phenotype appears to be endless, making the process of unraveling the genetics of allergy into a monumental task. Nevertheless, an awareness of the possible sources of the allergic reaction can aid in diagnosis and treatment of allergic conjunctivitis.
Allergy Activation
The first encounter with an allergen does not actually result in the signs and symptoms of allergy. Instead, the allergen binds to and is internalized by B cells, which in turn display antigenic peptides on their surface. The B cells then form conjugates with Th2 cells, and the resultant production of cytokines cause the allergen-bound B cells to differentiate into plasma cells. In an individual with a tendency toward a type-1 hypersensitivity response, immunoglobulin E (IgE) is secreted. IgE antibodies have a high affinity for the fragment crystallization (Fc) receptors on the surface of mast cells, and once they are bound, the mast cell is primed for the subsequent exposure to the allergen. During the second encounter with the allergen, the allergen cross-links with the IgE antibodies, triggering the mast cell to degranulate and release various allergic mediators. Every step of the priming and eventual allergic response is subject to environmental and genetic influences.
The allergic tendency — that is, to to overproduce IgE antibodies — is believed to be inherited. Therefore, parents' pollen allergies may manifest as a cat allergy in their children.7 Statistically significant relationships have been found between parental history of asthma and atopy and the incidence of these conditions in their children.8 Nevertheless, pinpointing the specific genes involved has proved difficult. Twin studies have suggested that upper and lower respiratory atopic symptoms are different manifestations of the same genes;7 asthma and rhinitis may also be connected.9 Studies with both identical and fraternal twins have shown differences in the prevalence of the various atopic disorders between the sexes.10 Twin studies have historically been vital to genetic studies; however, there are variables related to allergy that can complicate any conclusions. Twins are often born prematurely; this can correlate to both a lower incidence of allergy,11 as well as to a higher risk of lung damage and the development of asthma.12 Even so, some genes have been identified as being involved in the primary clinical phenotypes of atopic diseases.
The Role of GSTs
Glutathione-S-transferases (GSTs) are enzymes that use the products of oxidative stress as substrates, thus protecting cells from the damaging effects of reactive oxygen species (ROS). ROS are produced through the activation of mast cells, eosinophils, macrophages and neutrophils — all of which are involved in the pathogenesis of asthma and allergy. Each class of GST enzyme — mu, theta, and pi — preferentially metabolizes a specific ROS product.13
Although GSTs have been recognized as important for the detoxification of ROS for decades, their role in asthma and atopy is a more recent discovery. Various genes on chromosome 11q have shown associations with bronchial hyperresponsiveness and atopy; in particular, genetic variations of GSTs are common14,15 and influence the allergic response. One of the common polymorphisms of the pi-class GST gene (GSTP1) involves the substitution of valine for isoleucine at codon 105. Since codon 105 forms part of the active binding site, catalytic activity is diminished.16 This association between GSTP1 Val105/Val105 and atopy was first noticed in a study conducted with asthmatic subjects.17 These researchers noted that the GSTP1 Val105/Val105 mutation corresponded to a decreased risk of asthma and atopy; however, other researchers have found the opposite to be true, especially in the presence of ozone.14 This discrepancy could be the result of small study groups, which limit the ability to adequately study all the variants. Furthermore, other genetic and environmental interactions could influence the analysis. For example, the homozygous deletion of of the mu-class GST gene (GSTM1) prevents the production of the associated enzyme.15 This null genotype has been associated with decreased lung function; again, ozone levels intensify this effect.14 Therefore, subjects with a GSTP1 Val105/Val105 mutation and a GSTM1 homozygous deletion may have a higher risk of developing atopic diseases than subjects with a GSTP1 Val105/Val105 mutation and a normal GSTM1 expression. Nevertheless, the importance of GSTs for relieving oxidative stress is irrefutable.
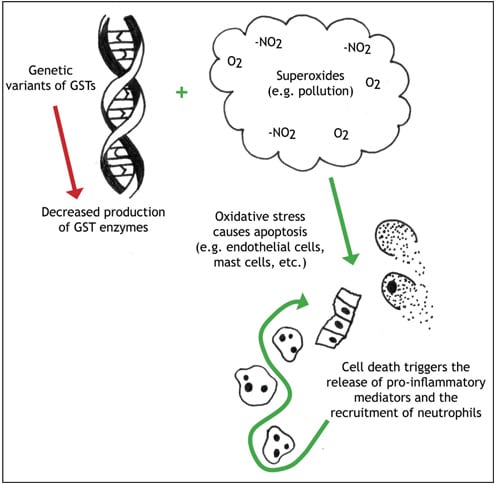
Linking genetics, pollution and allergic inflammation.
ROS and Eye Allergy
ROS are normal byproducts of cellular metabolism, and are important cell-signaling molecules. An overabundance of ROS, however, leads to apoptosis and inflammation. The pollution that typifies urban life — which involves high levels of ROS — may account for the rising incidence of urban allergy. When diesel exhaust hydrocarbons and nitrous oxide interact with sunlight, low-level ozone (O3) is produced. For some people, exposure to ozone and pollutants may prompt an allergic response that subsides over time. This has been demonstrated in laboratory mice (perhaps descended from the city mouse in Aesop's fable): upon exposure to diesel exhaust, their allergic response to ovalbumin is enhanced, but this reaction subsides with continuous exposure.18 Diesel exhaust particles may also enhance IgE production, and in allergy-prone individuals, diesel exhaust can induce sensitization to antigens that had never previously elicited an allergic response.19 Diesel exhaust particles may also lower the threshold at which an individual manifests the signs and symptoms of rhinitis, and as with the mice, especially so during high-dose, short-term exposure.20 Unfortunately for city mice and human urbanites, the doomsday warnings of pollution and genetics and the signs and symptoms of allergy do not end here.
There are several different classes of Fc receptors that are found on the surface of mast cells, including FcεR, which has a high affinity for IgE molecules. It is composed of three subunits; the ß chain is associated with enhancing the cell-surface expression of the receptor. Like the GST-mediating genes, FcεR is found on chromosome 11q, and polymorphisms in the FcεR gene are believed to be involved in the presentation of allergy.21 Several single nucleotide polymorphisms in the coding region result in amino acid substitution; however, their association with the presentation of allergy has not been consistent. It has therefore been hypothesized that the noncoding promoter region of FcεRIß may be the primary connection between the receptor and allergy. These polymorphisms of the mast cell Fc receptors can affect the number of receptors available to bind IgE antibodies.22 This increase in primed mast cells likely corresponds to a higher availability of binding sites for allergens and therefore the possibility for a more severe allergic reaction.
The genes that are potentially involved in the development of allergy are as numerous as the co-involved environmental factors. Identifying these genes is important for understanding allergic disorders and for the eventual development of gene therapies — and perhaps even preventative measures against allergy.
Tailoring Treatments
Patients who understand the causes of their allergies may be able to avoid the offending allergens and, when avoidance is impossible, treat their signs and symptoms with medications prescribed by their clinician. Topical medications are frequently favored, as they do not cause ocular drying and have a low incidence of adverse side effects. Olopatadine 0.1% and olopatadine 0.2% (Patanol, Alcon) are the most commonly prescribed dual-action antihistamine/mast cell stabilizers; olopatadine 0.2% has the added benefit of once-daily dosing. Epinastine 0.05% (Elestat, Allergan) is the second most widely prescribed dual-action agent. These medications have impressive efficacy and safety records. Topical corticosteroids, such as loteprednol 0.2% (Zylet, Bausch & Lomb) can be used in patients with more severe signs and symptoms. Just as the allergic reaction is specific to the genetic makeup and environment of the individual, treatment for the signs and symptoms of allergic conjunctivitis must be tailored to the needs of the individual. OM
References
1. Abelson MB, Smith L, Chapin M. Ocular allergic disease: mechanisms, disease sub-types, treatment. Ocular Surface. 2003;1:38-60.
2. Ayatollahi M, Geramizadeh B, Yazdani M, Azarpira N. Effect of immunoregulatory cytokines on successful pregnancy depends on the control of graft rejection mechanisms. Transplantation Proc. 2007;39:244-245.
3. Holland D, Bretscher P, Russell AS. Immunologic and inflammatory responses during pregnancy. J Clin Lab Immunol. 1984;14:177-179.
4. Astle S, Newton R, Thronton S, Vatish M, Slater DM. Expression and regulation of prostaglandin E synthase isoforms in human myometrium with labour. Mo Hu Reprod. 2007;13:69-75.
5. Viinanen A, Munhbayarlah S, Zevgee T, et al. The protective effect of rural living against atopy in Mongolia. Allergy. 2007;62:272-280.
6. Grant R, Bowen S, McLean DE, Berman D, Redlener K, Redlener I. Asthma among homeless children in New York City: an update. Am J Pub Health. 2007;97:448-450.
7. Thomsen SF, Ulrik CS, Kyvik KO, Ferreira MAR, Backer V. Multivariate genetic analysis of atopy phenotypes in a selected sample of twins. Clin Exp Allergy. 2006;36:1382-1390.
8. Alford SH, Zoratti E, Peterson EL, et al. Parental history of atopic disease: Disease pattern and risk of pediatric atopy in offspring. J Allergy Clin Immunol. 2004;114:1046-1050.
9. Guerra S, Sherrill DL, Martinez FD, Barbee RA. Rhinitis as an independent risk factor for adult-onset asthma. J Allergy Clin Immunol. 2002;109:419-425.
10. Lichenstein P, Svartengren M. Genes, environments, and sex: factors of importance in atopic diseases in 79-year-old Swedish twins. Allergy. 1997; 52:1079-1086.
11. Siltanen M, Kajossari M, Pohjavuori M, Savilahti S. Prematurity at birth reduces the long-term risk of atopy. J Allergy Clin Immunol. 2001;107:229-34.
12. Halvorsen T, Skadberg BT, Røksund OD, Carlsen KH, Bakke P. Pulmonary outcome in adolescents of extreme preterm birth: a regional cohort study. Acta Pædiatr. 2004;93:1294-1300.
13. Hayes JD, Strange RC. Potential contribution of the glutathione S-transferase supergene family to resistance to oxidative stress. Free Rad Res Commun. 1995;22:193-207.
14. Romieu I, Ramirez-Aguilar M, Sienra-Monge JJ, et al. GSTM1 and GSTP1 and respiratory health in asthmatic children exposed to ozone. Eur Respir J. 2006; 28:953-959.
15. Gilliland FD, Lin Y, Dubeau L, et al. Effects of glutathione S-transferase M1, maternal smoking during pregnancy and environmental and environmental tobacco smoke on asthma and wheezing in children. Am J Respir Crit Care Med. 2002;166:457-463.
16. Johansson AS, Stenberg G, Widersten M, Mannervik B. Structure-activity relationships and thermal stability of human glutathione transferase P1-1 governed by the H-site residue 105. J Mol Biol. 1998;278:687-698.
17. Fryer AA, Bianco A, Hepple M, Jones PW, Strange RC, Spiteri MA. Polymorphism at the glutathione S-transferase GSTP1 locus. Am J Respir Crit Care Med. 2000;161:1437-1442.
18. Matsumoto A, Hiramatsu K, Li Y, et al. Repeated exposure to low-dose diesel exhaust after allergen challenge exaggerates asthmatic responses in mice. Clin Immunol. 2006;121:227-235.
19. Diaz-Sanchez D, Garcia MP, Wang M, Jyrala M, Saxon A. Nasal challenge with diesel exhaust particles can induce sensitization to a neoallergen in the human mucosa. J Allergy Clin Immunol 1999;104:1183-1188.
20. Diaz-Sanchez D, Garcia MP, Saxon A. Diesel exhaust particles directly induce activated mast cells to degranulate and increase histamine levels and symptom severity. J Allergy Clin Immunol. 2000;106:1140-146.
21. Sandford AJ, Shirakawa T, Moffatt MF, et al. Localization of atopy and ε subunit of the high-affinity IgE receptor (FcεRI) on chromosome 11q. Lancet. 1993;341:332-334.
22. Nishiyama C, Azizawa Y, Nishyama M, et al. Polymorphisms in the FcεRIε promoter region affecting transcription activity: A possible promoter-dependent mechanism for association between FcεRIε and atopy. J Immunol. 2004;173:6458-64.
| Mark B. Abelson, M.D., an associate clinical professor of ophthalmology at Harvard Medical School and senior clinical scientist at Schepens Eye Research Institute, consults in ophthalmic pharmaceuticals. Lauren Lilyestrom is a medical writer at ORA Clinical Research & Development in North Andover, Mass. |

|